










De la EEA INTA Pergamino
Matías Dominguez1, 2; Julio Gonzalez1
1.Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Pergamino, Mejoramiento Genético de Girasol - 2.Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
RESUMEN
El primer paso en un programa de desarrollo de cultivares es formar una población con variabilidad genética para los caracteres de interés. La selección de progenies provenientes de poblaciones constituye una valiosa herramienta para el mejoramiento genético. El germoplasma estudiado proviene del cruzamiento del compuesto Bulk Vert, formado por la recombinación de líneas endocriadas de buen comportamiento frente a Verticillium dahliae, por el compuesto Bulk AO originado por la recombinación de líneas endocriadas de alto contenido de ácido oleico. Constituyendo la población Bulk Vert x Bulk AO de la EEA INTA Pergamino de citoplasma mantenedor de la fertilidad compuesta por diversas fuentes de germoplasma de origen local y exótico. El objetivo principal de la mejora de esta población es derivar líneas endocriadas para la obtención de híbridos y los objetivos específicos fueron evaluar la heredabilidad y las asociaciones entre caracteres de interés agronómico en familias S0:1 de la población Bulk Vert x Bulk AO de la EEA Pergamino, el avance genético esperado por selección, determinar la influencia de los caracteres estudiados sobre el rendimiento y evaluar el uso de índices de selección para el mejoramiento de la población.
Los caracteres evaluados fueron contenido de aceite, altura de planta, días para floración, diámetro del tallo, diámetro del capítulo, número de aquenios/capítulo, peso de 100 aquenios y rendimiento de aquenios por planta. Los resultados obtenidos muestran la variabilidad existente en la población, observándose diferencias entre familias para todos los caracteres. El carácter número de aquenios por capítulo debe ser considerado como criterio de selección para la obtención de genotipos con mayor rendimiento. La utilización de índices de selección en dicha población lograría obtener respuestas mayores a las esperadas por selección de caracteres individuales. La aplicación de estrategias de selección sobre esta población permitiría lograr avances en el mejoramiento de caracteres relacionados con rendimiento de aquenio y contenido de aceite.
INTRODUCCIÓN
El primer paso en un programa de desarrollo de cultivares es formar una población con variabilidad genética para los caracteres de interés. La selección de progenies provenientes de poblaciones constituye una valiosa herramienta para el mejoramiento genético.
Las poblaciones para utilizarse para la obtención de cultiva- res pueden formarse desde un cruzamiento biparental o ser más complejas por involucrar un número mayor de parentales. Una estrategia muy utilizada en mejoramiento genético de cultivos es la selección recurrente que por medio de sucesivos ciclos de selección permite aumentar la frecuencia de alelos favorables y obtener genotipos superiores. Implica la evaluación de progenies, la selección y recombinación de las superiores para generar el siguiente ciclo de la población.
La variabilidad observada en una población está determinada por el genotipo de los individuos y por el ambiente que influye en la expresión del carácter. La heredabilidad en sentido amplio indica qué proporción de la variabilidad total es de origen genético (Allard, 2010).

El conocimiento de la heredabilidad permite elegir la estrategia más adecuada para mejorar un carácter determinado y predecir la ganancia genética por selección (Lamkey y Hallauer, 1987) que está directamente relacionada con la proporción de variancia genética de tipo aditivo (estimada por la heredabilidad en sentido estricto, componente de la heredabilidad en sentido amplio). El estudio de las asociaciones entre caracteres constituye una valiosa herramienta a la hora de decidir la estrategia de selección para aplicar y conocer los efectos indirectos de la selección sobre otros caracteres, para lo cual la estimación de las correlaciones genotípicas entre ellos puede proveer la información que aporte a ese objetivo. El análisis de coeficiente de sendero permite descomponer los coeficientes de correlación entre caracteres considerados en un diagrama de relaciones en efectos directos e indirectos e identificar las variables que tienen mayor influencia sobre el carácter que se pretende mejorar (Wright, 1923; Li, 1975). Los índices de selección permiten mejorar para varios caracteres simultáneamente. Existen diferentes tipos de índices y todos tienen como objetivo predecir el mérito genético de los genotipos para seleccionar, maximizar la ganancia genética por selección y proveer al mejorador de un criterio para evaluar y seleccionar varios caracteres simultáneamente (Cerón Rojas y Crossa, 2018).
Ortegón y Escobedo (1995) en un estudio sobre cinco líneas de girasol encontraron baja variabilidad genética y heredabilidad para los caracteres de rendimiento de grano y aceite. Goksksoy et al. (2002) informaron altos valores de heredabilidad para el diámetro de capítulo, el peso de 1000 aquenios y el rendimiento de aquenios por planta. Mishra et al. (2003) encontraron los mayores valores de heredabilidad para la altura de planta, peso de 100 aquenios y número de hojas por planta. Baez et al. (1988) encontraron qué peso de 1000 aquenios tuvo alta y positiva correlación genética con rendimiento en kg de aceite por hectárea. Por su parte, Sarno et al. (1992) encontraron alta asociación entre peso de 1000 aquenios y rendimiento. Chikkadevaiah et al. (2002) informaron asociación positiva entre rendimiento y peso de aquenio, ciclo a floración y rendimiento de aceite. En análisis de coeficiente de sendero en girasol Tahir et al. (2019) obtuvieron efectos directos altos y positivos de la altura de planta y el diámetro de capítulo sobre el rendimiento de aquenios por planta. Dusanic et al. (2004) y Hladni et al. (2010) informaron efectos directos positivos y al- tos del número de aquenio y del peso de 1000 aquenios sobre el rendimiento de aquenios por planta.
La verticilliosis, causada por el hongo Verticillium dahliae, es una enfermedad de gran incidencia económica en el cultivo de girasol en la Argentina, puede causar pérdidas de rendimiento de hasta 73% en cultivos muy afectados (Bertero de Romano et al., 1994; Pereyra et al., 1999). Por un lado, la búsqueda de líneas que reúnan resistencia a esta enfermedad, buen rendimiento de grano y alto contenido de aceite es uno de los máximos desafíos del mejoramiento del cultivo. Por otro lado, la salida del mercado de los aceites sometidos a procesos de hidrogenación, por el efecto perjudicial para la salud de los alimentos que contienen grasas trans, ofrece a Argentina (uno de los principales países productores de aceite de girasol alto oleico) un importante nicho de demanda. Este aceite combina su alta estabilidad con su efecto beneficioso en la alimentación. Por estas razones, la población que se analiza proviene del cruzamiento entre dos compuestos genéticos que tienen por un lado buen comportamiento frente a Verticillium dahliae y por otro lado altos porcentajes de ácido oleico en su aceite. Ambos compuestos genéticos son de citoplasma mantenedor de la fertilidad y la evaluación de caracteres per se en lugar de aptitudes combinatorias que se realizó en este trabajo, se debe a que en poblaciones mantenedoras se desea mejorar la performance de las líneas para tener un mayor rendimiento en lotes de producción de semilla híbrida y por lo tanto disminuir el costo de producción del híbrido.
El objetivo principal de la mejora de esta población es derivar líneas endocriadas para la obtención de híbridos. La caracterización de estas fuentes de recursos genéticos de INTA conforma un aporte de importancia disponible para el mejoramiento del cultivo de girasol.
Los objetivos específicos del trabajo fueron evaluar la heredabilidad y las correlaciones genotípicas entre caracteres de interés agronómico en familias S0:1 de la población Bulk Vert x Bulk AO de la EEA Pergamino, el avance genético esperado por selección, determinar la influencia de los caracteres estudiados sobre el rendimiento y evaluar el uso de índices de selección para el mejoramiento de la población.
MATERIALES Y MÉTODOS
El germoplasma estudiado proviene del cruzamiento del compuesto Bulk Vert, formado por la recombinación de líneas de buen comportamiento frente a Verticillium dahliae, cruzado por el compuesto Bulk AO originado por la recombinación de líneas de alto contenido de ácido oleico. La población Bulk Vert x Bulk AO de la EEA INTA Pergamino resultante compren- de diversas fuentes de germoplasma de origen local y exótico con citoplasma mantenedor de la fertilidad. La población se condujo en dos ciclos de libre recombinación en lote aislado iniciándose en 2015/16 un ciclo de selección recurrente.

En la campaña 2015/16 se autofecundaron plantas en el compuesto Bulk Vert x Bulk AO. En 2016/17 se sembraron 67 familias S0:1 en la EEA Pergamino (Buenos Aires, Argentina, 33° 56’ S, 60° 33’ O) en un diseño de bloques completos al azar con tres repeticiones. La parcela experimental fue de un surco de 6 metros de largo por 0,52 metros entre parcelas. La fecha de siembra fue el 17 de noviembre de 2016 y el suelo donde se realizó el experimento fue un Argiudol típico de la serie Pergamino. Se midió contenido porcentual de aceite (AC) por la técnica de resonancia magnética nuclear de 1H resuelta en el tiempo (RMN) con un equipo Oxford MQ5 calibrado por extrac ción con solvente, altura de planta (cm) (ALT), días a floración (DAF), diámetro del tallo a 20 cm por debajo del capítulo (cm) (DT), diámetro del capítulo (cm) (DC), número de aquenios/ca pítulo (NAC), peso de 100 aquenios (g) (P100) y el rendimiento de aquenios por planta (g) (RAP). La altura de planta fue tomada en R5.5, el diámetro del tallo y de capítulo fue registrado en R9 (Scheneiter y Miller, 1981). Las mediciones se realizaron sobre cinco plantas de cada parcela a una distribución espacial y densidad adecuada con excepción de días a floración en la que se tomó un dato general de la parcela.
La estimación de los componentes de variancia fue realizada por REML (Restricted Maximum Likelihood) utilizando modelos lineales mixtos a través del paquete lme4 de R (Bates et al., 2015; R Core Team, 2019). La significancia de los componentes de variancia fue evaluada a partir de los valores de la probabilidad asociada obtenidos por la aproximación de Sat- terthwaite utilizando el paquete lmerTest de R (Kuznetsova et al., 2017; Luke, 2017).
El modelo lineal mixto utilizado para cada variable analizada fue:Y = µ + P + F + R + ε
El cálculo de los componentes de varianza genética fue realizado según Hallauer et al. (2010) para el tipo de familia S0:1. Debido al tipo de familias, solo es posible obtener una estimación sesgada de la heredabilidad en sentido estricto, debido a la presencia eventual de un cuarto de la dominancia mediante la resolución de:
h2 = VA’/ [VA’ + Vee/r]
Donde VA’ es la variancia aditiva sobreestimada por la presencia de un cuarto de la variancia de la dominancia, r es el número de repeticiones y Vee es la variancia del error experimental. El componente del denominador [VA’ + Vee/r] corresponde a la variancia fenotípica.
La predicción de la respuesta a la selección para cada varia ble se calculó según Falconer y Mackay (1996) a través de la fórmula:
Respuesta a la selección= k * σp * h2
k= intensidad de selección estandarizada (5% k=2,06)
σp= desviación estándar fenotípica
h2= heredabilidad en sentido estricto
También se obtuvieron las respuestas por ciclo de selección expresadas en términos porcentuales respecto a la media poblacional para compararlas con las obtenibles en otras poblaciones o entre variables (Mariotti y Collavino, 2015).
Las correlaciones genotípicas entre variables fueron calculadas a partir de las medias de las parcelas, y se agregó la variable días a floración debido a que se tenían solamente datos a nivel parcela. Estas fueron obtenidas por medio del paquete Lavaan de R (Rosseel, 2012) realizando un análisis de la covarianza.
Se realizó un análisis de sendero para descomponer en efectos directos e indirectos las correlaciones de las variables aceite, altura de planta, diámetro del tallo, diámetro del capítulo, número de aquenios/capítulo y peso de 100 aquenios con el rendimiento de aquenios por planta. Los coeficientes de sendero fueron obtenidos a través del paquete Lavaan de R (Rosseel, 2012) con base en las correlaciones genéticas y el método de estimación de los parámetros utilizado fue el de máxima verosimilitud.
A partir del análisis de coeficiente de sendero se seleccionaron las variables para conformar los índices de selección para el mejoramiento del rendimiento de aquenios por planta. Las variables consideradas fueron: diámetro del tallo, diámetro del capítulo, número de aquenios/capítulo y peso de 100 aquenios. Se confeccionaron dos índices de selección sin restricciones: Base Linear Phenotypic Selection Index (BLPSI), Eigen Selection Index Method (ESIM) y un índice de selección con restricciones: Restricted Linear Phenotypic Selection Index (RLPSI). Los índices fueron construidos con el software Rindsel (Pacheco et al., 2018). En el índice RLPSI se restringieron las variables diámetro de capítulo y diámetro de tallo. Los índices fueron obtenidos a partir de los datos de planta individual. Se determinó la respuesta absoluta por selección de cada carácter con una intensidad de selección del 5%, la precisión de selección y la heredabilidad de cada índice.
RESULTADOS
En la tabla 1 se presenta el análisis de varianza realizado por REML para la población Bulk Vert x Bulk AO observándo-se diferencias significativas entre familias S0:1para todos los caracteres. Se aprecia mayor variabilidad en rendimiento de aquenios por planta y número de aquenios por capítulo. Los valores más bajos de coeficiente de variación se dieron en contenido de aceite y altura de planta. Los más altos valores de heredabilidad se obtuvieron en contenido de aceite, peso de 100 aquenios y altura de planta y el menor en rendimiento de aquenios por planta.

Referencias: AC = aceite, DC = diámetro de capítulo, DT = diámetro de tallo, ALT = altura de planta, NAC = número de aquenios/capítulo,
P100= peso de 100 aquenios, RAP= rendimiento de aquenios por planta, * = Valor de probabilidad asociada menor a 0,05. Elaboración propia.
En la tabla 2 se muestran las correlaciones genéticas para rendimiento de aquenios por planta, contenido de aceite, altura de planta, diámetro de capítulo, diámetro de tallo, número de aquenios por capítulo, peso de 100 aquenios y días a floración en la población Bulk Vert x Bulk AO.

Referencias: RAP = rendimiento de aquenios por planta, AC = aceite, ALT = altura de planta, DC = diámetro de capítulo, DT = diámetro de tallo, NAC = número de aquenios/capítulo, P100= peso de 100 aquenios, DAF = Días a floración, * = Valor de probabilidad asociada menor a 0,05.
Elaboración propia.
Altura de planta tuvo asociación genética negativa con diámetro de capítulo y positiva con rendimiento de aquenios por planta y días a floración. En cambio, diámetro de capítulo mostró coeficientes de correlación positiva con diámetro de tallo, número de aquenios por capítulo, peso de 100 aquenios y rendimiento de aquenios por planta. El diámetro de tallo se asoció positivamente con número de aquenios por capítulo, peso de 100 aquenios y rendimiento de aquenios por planta. El número de aquenios por capítulo tuvo correlación positiva con altura, peso de 100 aquenios y rendimiento de aquenios por planta y correlación negativa con días a floración. Este úl- timo carácter correlacionó negativamente con rendimiento de aquenios por planta, aceite y peso de 100 aquenios.
En la tabla 3 se presenta la respuesta a la selección en la población Bulk Vert x Bulk AO en términos absolutos y porcentuales para aceite, diámetro de tallo, diámetro de capítulo, al- tura de planta, número de aquenios por capítulo, peso de 100 aquenios y rendimiento de aquenios por planta.
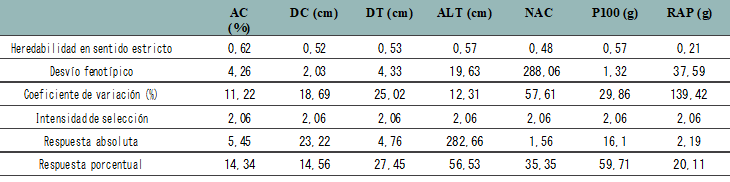
Referencias: AC = aceite, DC = diámetro de capítulo, DT = diámetro de tallo, ALT = altura de planta, NAC = número de aquenios/capítulo,
P100= peso de 100 aquenios, RAP= rendimiento de aquenios por planta. Elaboración propia
La tabla 4 muestra los efectos directos e indirectos obtenidos a partir del análisis de sendero de las variables aceite, altura de planta, diámetro del tallo, diámetro de capítulo, número de aquenios/capítulo y peso de 100 aquenios (g) sobre el rendimiento de aquenios por planta. Los efectos que superan en valor absoluto al efecto residual son los que influyen significativamente sobre el rendimiento (se indican en negrita en la tabla 4). El efecto directo del número de aquenios por capítulo, el efecto indirecto del diámetro de capítulo vía número de aquenios y el efecto indirecto del diámetro de tallo vía el número de aquenios son los de mayor importancia para el rendimiento por planta.

Coeficiente de determinación del modelo (R2) = 0,73. Efecto residual= 0,27. Referencias: AC = aceite, ALT = altura de planta, DC = diá-
metro de capítulo, DT = diámetro de tallo, NAC = número de aquenios/capítulo, P100= peso de 100 aquenios, DAF = Días a floración, r = correlación genética entre cada variable independiente con la variable dependiente (rendimiento de aquenios por planta).
Elaboración propia.
La tabla 5 muestra las respuestas absolutas por selección (intensidad de selección de % 5) para cada uno de los caracteres que componen los índices. Además, se presenta la pre- cisión de selección del índice, obtenida como la correlación entre el valor de mejora y el índice, y la heredabilidad de cada índice. El índice RLPSI es el que presenta la mayor respuesta para el carácter número de aquenios por capítulo, una elevada correlación con el valor de mejora y la mayor heredabilidad de los tres índices obtenidos.
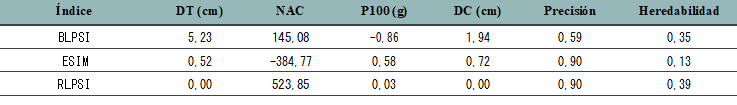
Referencias: BLPSI= Base Linear Phenotypic Selection Index, ESIM= Eigen Selection Index Method, RLPSI= Restricted Linear Phenotypic
Selection Index, DT = diámetro de tallo, NAC = número de aquenios/capítulo, P100= peso de 100 aquenios, DC = diámetro de capítulo. Elaboración propia
Por una parte, las condiciones climáticas (temperatura, precipitaciones y heliofanía) durante el ciclo del experimento fue- ron similares a las medias de los últimos veinte años, a excepción del mes de diciembre que tuvo precipitaciones mayores a la media. Por otra parte, la heliofanía acumulada mensual para los meses de febrero y marzo fueron menores a las medias pudiendo influir negativamente en la fase de llenado de las familias de ciclo más largo.
DISCUSIÓN Y CONCLUSIONES
Por una parte, la estimación de heredabilidad del contenido de aceite resultó similar a las reportadas por Waseem et al. (2004) y González et al. (2008) y la de rendimiento de aquenios por planta fue diferente a las de Ortegón y Escobedo (1995). Por otra parte, estos últimos autores informaron baja heredabilidad para diámetro de capítulo. Estas diferencias pueden deberse al número reducido de genotipos y a que la población evaluada por los autores fue de líneas endocriadas. Las heredabilidades para peso de 100 aquenios, altura de planta y diámetro de capítulo fueron similares a las informadas por Goksksoy et al. (2002) y Mishra et al. (2003).
Las asociaciones genotípicas positivas entre peso de 100 aquenios con número de aquenios por capítulo y entre peso de 100 aquenios y rendimiento de aquenios por planta se asemejan a lo publicado por otros autores como Baez et al. (1988), Chikkadevaiah et al. (2002) y Lakshminarayana et al. (2004).
El contenido de aceite tuvo asociación negativa con días de siembra a floración. Baez et al. (1988) también encontraron asociaciones negativas entre estas variables. En cambio Fick et al. (1974) encontraron correlaciones positivas y ausencia de correlación entre esas variables tanto al analizar cultivares de polinización abierta como cultivares híbridos. Otros autores como Putt (1943) y Ross (1939) no encontraron asociaciones entre ambos caracteres.
A diferencia de Cecconi et al. (1987) y Hladni et al. (2004) que obtuvieron asociaciones genéticas positivas entre la altura de planta y el diámetro de capítulo, la correlación entre estos caracteres para la población estudiada fue negativa. Podría deberse a una competencia por fotosintatos entre la elongación de entrenudos y el crecimiento del capítulo (Aguirrezabal et al., 1996). Desde el punto de vista de la selección, esta relación negativa resultaría favorable para lograr un ideotipo de planta de menor altura para evitar el vuelco y quebrado de tallo y un mayor diámetro de capítulo por su relación positiva con el rendimiento.
Las diferencias encontradas en las correlaciones genotípicas entre caracteres con otros autores pueden deberse a que la estimación de estas está fuertemente influenciada por las frecuencias génicas de cada población, existiendo además errores de muestreo que disminuirían su precisión. Asimismo, como una de las causas de las correlaciones genotípicas entre caracteres es el ligamiento, es probable que entre poblaciones se presenten correlaciones diferentes dependiendo del tipo de población, los ciclos de recombinación y el desequilibrio de ligamiento. Esto resalta la importancia de realizar una caracterización de cada población en particular que se pretende utilizar para la derivación de líneas.

El análisis de coeficientes de sendero permitió descomponer las correlaciones genéticas entre las variables estudiadas, mostrando que el efecto directo del número de aquenios por capítulo y los efectos indirectos del diámetro del capítulo y del diámetro de tallo por medio del número de aquenios fueron positivos y significativos sobre el rendimiento de aquenios por planta. Estos resultados fueron acordes a los obtenidos por Hladni et al. (2008), quienes identificaron efectos directos positivos del número de aquenios por capítulo y efectos indirectos del diámetro de tallo y del diámetro de capítulo vía el número de aquenios por capítulo. También obtuvieron un efecto directo negativo no significativo del contenido de aceite sobre el rendimiento.
La altura de planta y el peso de 100 aquenios evidenciaron efectos directos positivos, pero no significativos sobre el rendimiento, similares conclusiones obtuvieron Yasin y Singh (2010) con respecto a la influencia de la altura de planta y Kaya et al. (2009) con el peso de 100 aquenios sobre el rendimiento. El efecto directo negativo de los días a floración sobre el rendimiento obtenido fue similar a lo informado por Farhatullah et al. (1996). En cambio, Riaz et al. (2019) registra- ron efectos directos negativos de la altura de planta y efectos directos significativamente positivos del diámetro de capítulo sobre el rendimiento de aquenios. Estos autores también obtuvieron efecto directo negativo no significativo del contenido de aceite sobre el rendimiento, permitiendo mejorar simultánea- mente por ambos caracteres sin que la mejora en uno implique una disminución en la performance del otro.
Las diferencias obtenidas en cada caso con otros autores deben ser tenidas en cuenta considerando que los resultados obtenidos en este trabajo se atañen a la población particular y al ambiente específico donde se realizó el experimento.
Los índices de selección evaluados mostraron ganancias genéticas esperadas diferentes para los caracteres que mayor efecto tienen sobre el rendimiento. Si bien al momento de tomar la decisión sobre cuál índice es conveniente usar se debe tener en cuenta en primera instancia la precisión de selección (correlación entre el índice y el valor de mejora), autores como Cerón Rojas et al. (2008) y Smith et al. (1981) resaltan la importancia de tener en cuenta la ganancia genética esperada por selección que se obtiene con cada índice y considerar si se ajustan a los objetivos del programa de mejoramiento. En el caso del índice ESIM la precisión de selección fue máxima, pero la ganancia genética esperada del carácter número de aquenios por capítulo resultó negativa, en cambio el índice BLPSI registró menor precisión de selección, pero una mayor ganancia genética para el número de aquenios por capítulo. El índice restringido RLP- SI obtuvo una máxima precisión de selección, fue el de mayor ganancia genética esperada para el número de aquenios por capítulo y el de mayor heredabilidad. Los resultados obtenidos indican que para la población estudiada el índice RLPSI es la mejor alternativa entre los índices evaluados.
De acuerdo a los análisis realizados en el presente trabajo, el empleo de estrategias de selección sobre la población Bulk Vert x Bulk AO de INTA Pergamino permitirá lograr avances en el mejoramiento de caracteres relacionados con rendimiento de aquenios y el contenido de aceite. A partir del análisis de coeficientes de sendero se desprende que el carácter número de aquenios por capítulo debe ser considerado como criterio de selección para la obtención de genotipos con mayor rendimiento. La utilización de índices de selección es una herramienta de utilidad para el mejoramiento de esta población, pudiendo ser implementados en estrategias de selección recurrente permitiendo obtener respuestas mayores a las esperadas por selección de caracteres individuales.
AGRADECIMIENTOS
Los autores agradecen a los doctores Guillermo Eyherabide y Guillermo Pratta por sus valiosas colaboraciones en los análisis genéticos y en la revisión del manuscrito, al grupo de Mejoramiento de girasol de INTA Pergamino y a los alumnos de la Escuela de Educación Agraria N.º 1 de Pergamino por la colaboración con el ensayo y toma de datos, y a la Lic. María Jose Beribe por su apoyo con los análisis estadísticos.
BIBLIOGRAFÍA
AGUIRREZABAL, L.; ORIOLI, G.; HERNANDEZ, L.; PEREYRA, V.; MIRAVE, J. 1996. Girasol: aspectos fisiológicos que explican el rendimiento. Ed. Unidad Integrada Balcarce. Balcarce, Argentina. 111 p.
AGUIRREZABAL, L.; MARTRE, P.; PEREYRA-IRUJO, G.; ECHARTE, M.; IZQUIERDO, N. 2015. Improving grain quality: ecophysiological and modeling tools to develop management and breeding strategies. En: SADRAS, V.; CAL- DERINI, D. (Eds.). Crop Physiology: Applications for genetic improvement and agronomy. Elsevier. 423-465 pp.
ALLARD, R. 2010. Principles of Plant Breeding. Ed. Wiley. India.
ANDRIANASOLO, F.; DEBAEKE, P.; CHAMPOLIVIER, L.; MAURY, P. 2016.
Analysis and modelling of the factors controlling seed oil concentration in sunflower: a review. OCL, 23(2) D206.
BAEZ, M.; LUDUEÑA, P.; SANGUINETTI, A. 1988. Correlaciones fenotípi- cas, genotípicas y ambientales entre caracteres de girasol y podredumbre del tallo (Macrophomina phaseoli (Maub.) Ashby). 12th International Sun- fIower Conference. Novi Sad, Yugoslavia. 11. 519-524.
BATES, D.; MAECHLER, M.; BOLKER, B.; WALKER, S. 2015. Fitting Linear Mixed-Effects Models Using lme4. Journal of Statistical Software. 67 (1), 1-48.
BERTERO DE ROMANO, A.; VAZQUEZ, A.; PIUBELLO, S.; SALA, C. 1994. Quantifying the relationship between Verticillium Wilt Intensity and yield loss of Sunflower (Helianthus annuus L.), and feasibility of utilizing the hy- podermic inoculation technique as a selection method. Helia, 17, 20. 49-53.
CECCONI, F.; PUGLIESI, C.; BARONCELLI, S.; ROCCA, M. 1987. Genetic analysis for some agronomical characters of a sunflower (Helianthus an- nuus L.) diallel cross. Helia, 10: 21-27.
CERÓN-ROJAS, J.; SAHAGÚN-CASTELLANOS, J.; CASTILLO-GONZÁLEZ, F.; SANTACRUZ-VARELA, A.; CROSSA, J. 2008. A Restricted Selection Index Method Based on Eigenanalysis. J. Agric. Biol. and Env. Statistics, 13: 440-457.
CERÓN-ROJAS, J.; CROSSA, J. 2018. Linear Selection Indices in Modern Plant Breeding. Ed. Springer open. México. 256 p.
CHIKKADEVAIAH; SUJATHA, H.L.; NANDINI. 2002. Correlation and path analysis in sunflower. Helia. 25: 109-118.
DUSANIC, N.; MIKLIC, V.; JOKSIMOVIC, J.; ATLAGIC, J. 2004. Path coefficient analysis of some yield components of sunflower. 16th International Sunflower Conference. Vol. ii. 531-537. Fargo, ND, EUA.
FARHATULLAH; E-AZAM, F.; KHALIL, I. 2006. Path Analysis of the Coeffi- cients of Sunflower (Helianthus annuus L.) Hybrids. Int. J. Agri. & Biol. 8: 621-625.
FICK, G.; ZIMMER, E.; ZIMMERMAN, D. 1974. Correlation of Seed Oil Con- tent in Sunflowers with Other Plant and Seed Characteristics. Crop Science Vol. 14. 755-757.
GOKSKSOY, A.; TURKEC, A.; TURAM, Z. 2002. Quantitative inheritance in sunflower (Helianthus annuus L.). Helia 37:131-140.
GONZÁLEZ, J.; MANCUSO, N.; LUDUEÑA P. 2008. Heredabilidad de compo-nentes de rendimiento en dos poblaciones de girasol de la EEA Pergamino. 17th International Sunflower Conference. Vol. ii. 567-570. Córdoba, España.
HALLAUER, A.; CARENA, J.; MIRANDA FILHO, J. 2010. Quantitative Genetics in Maize Breeding. Ed. Springer Nueva York. 663 p.
HLADNI, N.; SKORIC, D.; KRALJEVIC-BALALIC, M.; IVANOVIC, M.; SAKAC, Z.; JOVANOVIC, D. 2004. Correlation of yield components and seed yield per plant in sunflower (Helianthus annuus). 16th International Sunflower Confe- rence. Vol. ii. 491-496.
Fargo, ND, EUA. HLADNI, N.; JOSIC, S.; MIKLIC, V.; MIJIC, A.; PANKOVIC, D. 2008. Direct and indirect effects of morphophysiological traits on seed yield of sunflower (Helianthus annuus L.). 17th International Sunflower Conferen- ce. Vol. i: 393-397.Córdoba, España.
HLADNI, N.; JOSIC, S.; MIKLIC, V.; MIJIC, A.; SAFTIC-PANKOVIC, D.; SKORIC, D. 2010. Effect of morphological and physiological traits on seed yield and oil content in sunflower. Helia 39: 101-116.
KAYA, Y.; EVCI, G.; DURAK, S.; PEKCAN, V.; GUCER, T. 2009. Yield components affecting seed yield and their relationships in sunflower (Helianthus annuus L.). Pak. J. Bot. 41(5): 2261-2269.
KUZNETSOVA, A.; BROCKHOFF, P.; CHRISTENSEN, R. 2017. lmerTest Package: Tests in Linear Mixed Effects Models. Journal of Statistical Software, 82(13), 1-26. doi:10.18637/jss.v082.i13
LAKSHMINARAYANA, N.; SREEDHAR, N.; PRABAKAR, A. 2004. Correlation and path analysis in sunflower (Helianthus annuus L.) The Andra Agric. J. 51: 342-344.
LAMKEY, K.; HALLAUER, A. 1987. Heritability estimated from recurrent se- lection experiments in maize. Maydica xxxii, 61:78.
LI, C. 1975. Path Analysis – A Primer. Ed. The Boxwood Press (EE. UU.). 346 p.
LUKE, S.G. 2017. Evaluating significance in linear mixed-effects models in R. Behav Res 49: 1494.
MARIOTTI, J.; COLLAVINO, N. 2015. Los caracteres cuantitativos en la mejora genética de los cultivos. Orientación Gráfica Editora. Buenos Aires, Argentina. 325 p.
MISHRA, D.; ROY, D. 2003. Heritability estimates in dwarf population of sunflower (Helianthus annuus L.). Helia 39:37-42.
ORTEGÓN, A.; ESCOBEDO, A. 1995. Variabilidad genética, fenotípica y correlaciones entre componentes de rendimiento de líneas de girasol (He- lianthus annuus). Agronomía Maesoamericana 6:151-156.
PACHECO, A.; PEREZ, S.; ALVARADO, G.; CERÓN, J.; RODRIGUEZ, F.; CROSSA, J.; BURGUEÑO, J. 2018. RIndSel (Selection Index with R for Windows). Texcoco, México. Centro Internacional de Mejoramiento de Maíz y Trigo. (Disponible: https://data.cimmyt.org/dataverse/cimmytswdvn verificado: 05 de septiembre de 2019).
PEREYRA, V.; QUIROZ, F.; AGÜERO, M.; ESCANDE, A. 1999. Relación del rendimiento del girasol con la intensidad de síntomas provocados por Verti- cillium dahliae. x Jornadas Fitosanitarias Argentinas. Jujuy. 35 p.
PUTT, E. 1943. Association of seed yield and oil content with other characters in the sunflower. Sci. Agri. 23: 377-383.
R CORE TEAM. 2019. R: A language and environment for statistical com- puting. R Foundation for Statistical Computing, Vienna, Austria. (Disponible: https://www.R-project.org/ verificado: 02 de mayo de 2019).
RIAZ, A.; HAMMAD, M.; TAHIR, N.; RIZWAN, M.; FIAZ, S.; CHACHAR, S.; RAZZAQ, K.; RIAZ, B.; SADIA, H. 2019. Developing a Selection Criterion Using Correlation and Path Coefficient Analysis in Sunflower (Helianthus annuus L.). Helia 42: 85-99.
ROSS, A. 1939. Some morphological characters of Helianthus annuus L., and their relationship to the yield of seed and oil. Sci. Agr. 19: 372-379.
SARNO, R.; LETO, C.; CARRUBA, A.; CIBELLA, R. 1992. Correlation between some yield factors in sunflower (Helianthus annuus L.). 13th International Sunflower Conference. Pisa Italy Vol. 1: 366-389.
SCHNEITER, A.; MILLER, J. 1981. Description of Sunflower Growth Stages. Crop Science, 21, 901-903.
SMITH, S.; HALLAUER, A.; RUSSELL, W. 1981. Use of index selection in recurrent selection programs in maize. Euphytica 30, 611-618.
TAHIR, A.; IQBAL, M.; SAIF, R.; QADIR, M.; SULTANA, R. 2019. Correlation and Path Coefficient Analysis for Morphological and Biochemical Parame- ters in Sunflower (Helianthus annuus L.). Helia. 42: 61-72.
WASEEM, H.; SADAQAT, M.; SHAHIDA, H. 2004. Variability for agronomic traits in two random-mating sunflowers populations: means, variance com- ponents and heritabilities. Helia 41:99-112.
WRIGHT, S. 1923. The theory of path coefficients a reply to Niles´s criti- cism. Genetics vol. 8 n.o 3: 239-255.
YASIN, A.; SINGH, S. 2010. Correlation and path coefficient analysis in sunflower. Journal of Plant Breeding and Crop Science. Vol. 2(5). 129-133
















