











Por: Fernando H. Oreja, Juan I. Puente, Elba B. de la Fuente -Catedra de Cultivos Industriales de la Facultad de Agronomía de la Universidad de Buenos Aires
Amaranthus palmeri es una especie anual, de metabolismo C4 (Wang et al., 1992), originaria del desierto de Sonora y el valle inferior del Río Grande, ubicados en el sudoeste de Estados Unidos y noroeste de México (Sauer 1957; Ehleringer, 1983) (Figura 1). Una región que se caracteriza por su clima árido, suelos con poca disponibilidad de nutrientes y alta variación térmica.
En la Argentina, si bien fue registrada en materiales de herbarios hace unos 45 años, recién fue registrada por primera vez como maleza problemática en el año 2012 (Morichetti et al. 2012). A partir de ese momento se expandió a gran velocidad llegando a ocupar gran parte del área productiva (REM 2022) incluidos algunos países limítrofes.

Figura 1. Ubicación del Desierto de Sonora en Norteamérica.
El hecho de haber evolucionado en ambientes áridos y cálidos le permitió adquirir características únicas que la hicieron una excelente competidora por recursos frente a otras especies. Una de las características es su gran capacidad de capturar radiación solar a través de la orientación de sus hojas en forma perpendicular a los rayos solares (conocido como diaheliotropismo, Ehleringer y Forseth 1980). Otra es la disposición de las hojas en el tallo de manera tal que las hojas nuevas que van saliendo no sombrean a las anteriores. Esta eficiente captura de radiación junto a la alta eficiencia en el uso de esa radiación, incluso más alta que otras especies C4 (Ehleringer 1983), le permite alcanzar tasas de crecimiento muy altas (3 a 4 cm por día) sobre todo durante los primeros estadios fenológicos. Además, con restricciones hídricas, puede acumular solutos en las hojas que le permiten mantener los estomas abiertos realizando fotosíntesis (Ehleringer 1985). En lotes de soja de Arkansas, Estados Unidos, se han registrado pérdidas de hasta el 48% con unas 3 plantas por m2 (Klingaman y Oliver, 1994) y en lotes de maíz con 8 plantas por metro las pérdidas llegaban hasta el 74% (Massinga y Currie 2002).
Por otra parte, a diferencia de la mayoría de los yuyos colorados que son hermafroditas, esta planta es diclino-dioica, es decir que tiene individuos machos y hembras por separado. Esta particularidad le permite tener una alta variabilidad genética que no solo genera una gran diversidad morfológica entre individuos de la misma especie, sino que aumenta las chances de que aparezcan individuos con resistencia a herbicidas.
Desde el primer registro de un biotipo resistente a herbicidas en 1989 en Carolina del Sur, en el sudeste de Estados Unidos, hasta la fecha se registraron en promedio dos biotipos por año, llegando a la actualidad a 70 casos de esta especie con resistencia a nueve modos de acción distintos alrededor del mundo (Heap 2022). Lo que hace cada vez más difícil su manejo, especialmente en planteos basados únicamente en el uso de herbicidas.
Por ello es necesario elaborar y diseñar estrategias de manejo integrado que favorezcan la habilidad competitiva del cultivo frente a la maleza resulta de suma importancia para reducir la incidencia de esta especie, donde el uso de herbicidas es una herramienta importante pero no la única. El objetivo de este trabajo fue evaluar el efecto de la presencia del cultivo de soja con distintas estructuras de cultivo sobre el crecimiento y la fecundidad de esta especie.
Materiales y métodos
Diseño del experimento
El experimento se llevó a cabo en el campo experimental de la Cátedra de Cultivos Industriales de la Facultad de Agronomía de la Universidad de Buenos Aires (FAUBA), ubicado en la Ciudad Autónoma de Buenos Aires, Argentina (34° 35’ 32” S; 58° 29’ 16” O). Se realizó con un diseño en bloques completamente aleatorizados con 3 repeticiones donde los factores a evaluar fueron i) presencia de cultivo con dos niveles (con cultivo y sin cultivo), ii) densidad del cultivo (baja densidad, BD: 20 plantas m-2; alta densidad, AD: 35 plantas m-2) y iii) arreglo espacial (convencional, C y en grilla, G).

La siembra se realizó el 1 de diciembre, mediante una sembradora experimental de 4 cuerpos distanciados a 35 cm. En las parcelas en G se sembró dos veces con la mitad de la densidad correspondiente pasando dos veces, la segunda de forma perpendicular a la primera. La variedad utilizada fue Don Mario ® 3800, grupo de madurez 3.5. Luego de la emergencia de las plántulas de soja, se ralearon hasta obtener las densidades objetivo. A su vez, en subparcelas de 1 m-2 se sembraron semillas de A. palmeri en el entresurco, provenientes de establecimientos al Sur de Córdoba (Argentina). Luego, se raleo manualmente de manera de obtener 4 plantas por parcela (Figura 2).
Mediciones
Durante el experimento, se registró la fenología del cultivo y la radiación interceptada por el mismo de manera periódica. Para ello se utilizó un radiómetro, a las 12 am de días diáfanos, colocando el mismo en posición transversal a uno de los vértices de cada parcela. Se registró la temperatura media y las precipitaciones diarias de una estación meteorológica ubicada a unos 400 m.
Al final del experimento, se cosecharon las plantas de A. palmeri y un metro cuadrado de la parte área del cultivo del centro de cada parcela evitando las borduras. Luego se colocó en estufa hasta peso constante. Finalmente, se pesó la biomasa del cultivo de soja y los granos. En el caso de la maleza, se pesó la biomasa y se separó las semillas por medio de zarandas y un soplador para pesarlas.
Análisis de datos
Todas las variables analizadas en los tratamientos (rendimiento y biomasa del cultivo y biomasa y producción de semillas de la maleza) se sometieron a un análisis de varianza (ANVA) y posteriormente comparaciones múltiples (HSD Test de Tukey) para evaluar diferencias significativas (con un p<0,05) entre las medias de los tratamientos. Los análisis se realizaron con el paquete estadístico Infostat (Di Rienzo et al., 2013). Se evaluaron los supuestos del ANVA (homogeneidad de la varianza, observaciones independientes, distribución normal); en el caso de no cumplir con dichos supuestos se transformó dichas variables a la escala logarítmica en base dos.
Además, se estimaron tres medidas de competencia: complementariedad de recursos, habilidad competitiva, y severidad de competencia (Wilson, 1988). Por otra parte, se realizó un diagrama bivariado (Snaydon y Satorre, 1989) para interpretar las interacciones posibles entre la maleza y el cultivo. La complementariedad de recursos se midió a partir del “rendimiento total relativo” (RYT), el cual se calcula por la sumatoria de la producción (materia seca por unidad de área (MS)) relativa de biomasa del componente i (MSij / MSii) y del componente j (MSji / MSjj) (Satorre y Kruk, 2016).
RYTij= (MSij / MSii) + (MSji / MSjj) (ecuación 1)
La habilidad competitiva de la maleza y del cultivo, se cuantificó mediante el índice de “agresividad” (AG) (McGilchrist y Trenbath, 1971), que se calcula por la diferencia de la producción relativa de biomasa del componente i (MSij / MSii) y del componente j (MSji / MSjj).
AGij = (MSij / MSii) - (MSji / MSjj) (ecuación 2)
Además, la severidad de competencia (S), se estimó por separado a partir de la producción relativa de biomasa del cultivo y de la maleza (Satorre y Kruk, 2016).
Si = (MSij / MSii) y Sj = (MSji / MSjj) (ecuación 3)
Resultados y discusión
La presencia del cultivo de soja, independientemente de la estructura del cultivo, redujo significativamente el crecimiento en un 66% (Figura 3a) y la fecundidad de las plantas de A. palmeri en un 88% en promedio comparado con los tratamientos de la maleza creciendo sola (Figura 3b). Sin embargo, no se encontraron diferencias en la biomasa ni en la fecundidad producida por la maleza entre las distintas estructuras de cultivo testeadas.
Dado que no hubo deficiencias hídricas ni nutricionales, se podría afirmar que la disminución de la biomasa de A. palmeri en los tratamientos con cultivo fue debida principalmente a la competencia por radiación solar entre las plantas de soja y la maleza.
A los 30 días desde la emergencia del cultivo, alrededor de la etapa R1, todos los tratamientos excepto el BC (baja densidad) tuvieron valores de intercepción de radiación superiores a SC (Figura 4). Posiblemente esto se deba a cierto desfasaje fenológico entre la emergencia del cultivo y de la maleza, lo cual redujo la competencia interespecífica por radiación a favor del cultivo dado que pudo implantarse y establecerse antes que A. palmeri.
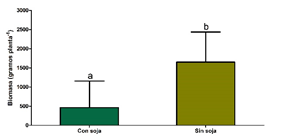
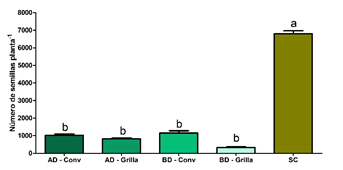
Figura 3. a) Biomasa aérea (g por planta) de A. palmeri con y sin cultivo de soja. b) Fecundidad de A. palmeri (semillas por planta) para los distintos tratamientos, alta densidad y siembra convencional (AD - Conv), alta densidad y siembra en grilla (AD - Grilla), baja densidad y siembra convencional (BD - Conv), baja densidad y siembra en grilla (BD - Grilla) y sin cultivo (SC).
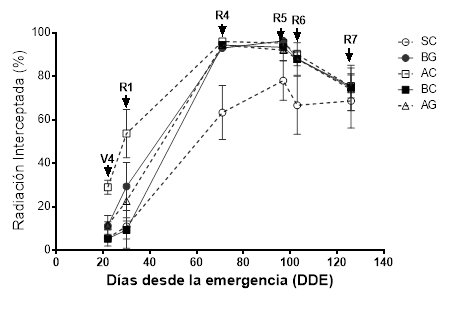
Figura 4. Radiación interceptada (%) por los tratamientos alta densidad y siembra convencional (AC), alta densidad y siembra en grilla (AG), baja densidad y siembra convencional (BC), baja densidad y siembra en grilla (BG) y sin cultivo (SC), en función del tiempo desde la emergencia del cultivo (DDE). Las flechas indican los estadios fenológicos del cultivo.
Las diferencias de intercepción de radiación se explican por la mayor habilidad competitiva y severidad de competencia que posee el cultivo en los diferentes tratamientos. Esto se refleja en los índices de agresividad (AG) y de rendimiento relativo del cultivo (RRc) estimados (Cuadro 1).
Sólo el tratamiento BC (baja densidad y siembra convencional) tuvo mayor rendimiento relativo o severidad de la maleza (RRm) y rendimiento relativo o severidad del cultivo (RRc) que el resto de los tratamientos. Los demás tratamientos no variaron respecto al BC (Cuadro 1), excepto del tratamiento BG (baja densidad y siembra en grilla) en el cual el rendimiento relativo total (RYT) fue menor a 1. Este resultado podría adjudicarse a una menor respuesta en el rendimiento en biomasa de la soja y a la incapacidad de la maleza de capturar los recursos no utilizados por el cultivo. En el caso del tratamiento BC tuvo valores cercanos a 2 indicando la existencia de complementariedad en el uso de los recursos entre las especies en estudio. En el caso de AC y AG se observan valores cercanos a 1 lo que sugiere la existencia de plena competencia entre las especies y en el tratamiento BG se ven valores inferiores a 1, posiblemente porque no compiten plenamente. Finalmente, el índice de agresividad (AG) determinó que el cultivo fue más competitivo que A. palmeri en todos los tratamientos (Cuadro 1).
Cuadro 1. Tabla de índices de competencia para los distintos tratamientos: alta densidad y siembra convencional (AC), alta densidad y siembra en grilla (AG), baja densidad y siembra convencional (BC) y baja densidad y siembra en grilla (BG).
| Tratamientos | Rendimiento relativo de la Maleza (RRm) | Rendimiento relativo del cultivo (RRc) | Rendimiento total relativo (RYT) | Agresividad (Agr) |
| AC | 0,13 | 0,93 | 1,06 | 0,80 |
| BC | 0,65 | 1,16 | 1,81 | 0,51 |
| BG | 0,11 | 0,60 | 0,72 | 0,49 |
| AG | 0,17 | 0,98 | 1,16 | 0,81 |
Las plantas de A. palmeri que crecieron con el cultivo de soja, además de tener menor biomasa que las plantas que crecieron sin el cultivo, tuvieron menor producción de semillas por planta (Figura 5). En la mayoría de los trabajos sobre competencia entre especies (Grime, 1979; Spitters, 1983; Tilman, 1988; Goldberg y Landa, 1991; Satorre y Snaydon, 1992; Wilson y Tilman, 1995) la variable respuesta medida es el crecimiento de las plantas (biomasa) que permite analizar cómo se particionan los recursos entre los componentes de la mezcla.
No obstante, la competencia puede afectar la fecundidad y/o la supervivencia (Aarssen y Keogh, 2002). De esta manera, se pueden determinar los efectos de la competencia no sólo durante el ciclo de la maleza sino a futuro con la descendencia (cantidad de semillas) que dejan las especies que están compitiendo.
El resultado encontrado podría estar mostrando una respuesta característica de lo que Weiner (2004) define como plasticidad aparente. La plasticidad aparente es aquella que ocurre cuando las plantas cambian su patrón de traslocación de fotoasimilados en respuesta a la competencia. De esta manera, cuando las plantas no compiten con sus vecinas por recursos, traslocan una gran proporción de fotoasimilados a estructuras reproductivas, pero cuando hay competencia con plantas vecinas, traslocan menos fotoasimilados a estructuras reproductivas y más a estructuras vegetativas como tallos y hojas. Diversos trabajos aplican este concepto, entre ellos Nurse y DiTommaso (2005), quienes observaron que la reducción en la producción de semillas de Abutilon theophrasti en presencia de maíz fue resultado de una menor cantidad de fotoasimilados destinados a reproducción. A su vez, Brainard et al. (2005) encontraron que plantas de Amaranthus powellii bajo sombreo, aumentan la partición de biomasa para el tallo y reducen la asignación a las semillas. También, Weiner (1988) encontró que A. powellii en competencia puede sufrir cambios en la asignación de biomasa priorizando estructuras que incrementen la habilidad competitiva (p.ej. tallo) y asignando menos a la reproducción.
Por lo tanto, las plantas de la maleza que crecieron con el cultivo además de tener menor cantidad de fotoasimilados disponibles, podrían haber particionado de manera distinta que aquellas que crecen sin la presencia del cultivo, dando como resultado una menor disponibilidad de fotoasimilados para estructuras reproductivas y en consecuencia una menor producción de semillas por planta.
Los valores de semillas por planta de A. palmeri encontrados se asemejan a los hallados por Massinga et al. (2001). Como en la gran mayoría de los trabajos, hubo gran variabilidad en dichos valores producto de las diversas condiciones en las que crecen las plantas tanto en competencia (Massinga et al., 2001) como en ausencia de esta (Keeley et al., 1987).
En general, no hubo diferencias significativas en la biomasa del cultivo entre los distintos tratamientos pese a la presencia de la maleza. Esto puede deberse, por un lado, al desfasaje fenológico entre el cultivo y la maleza en los primeros días después de emerger el mismo, y por otro lado la alta habilidad competitiva y severidad de competencia que tuvo el cultivo (Cuadro 1). Teniendo buena producción de biomasa e intercepción de la radiación por encima del 95% durante el periodo crítico del cultivo, la presencia de 4 plantas m-2 de A. palmeri tampoco generó reducciones en el rendimiento en grano (Figura 5).
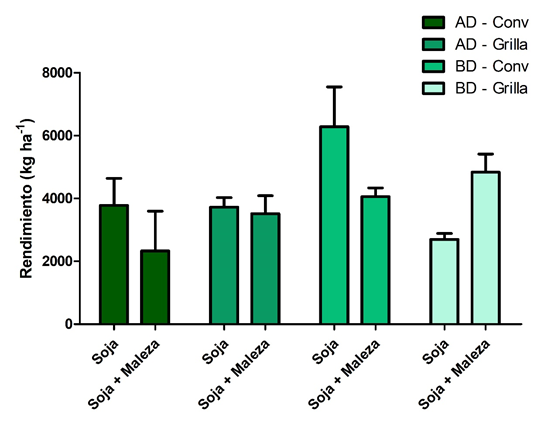
Figura 5. Rendimiento (kg/ha) del cultivo de soja para los distintos tratamientos, alta densidad y siembra convencional (AD - Conv), alta densidad y siembra en grilla (AD - Grilla), baja densidad y siembra convencional (BD - Conv), y baja densidad y siembra en grilla (BD - Grilla).
Conclusiones
- La soja redujo el crecimiento y la fecundidad de A. palmeri independientemente de la estructura del cultivo.
- La presencia de A. palmeri no afectó la biomasa ni el rendimiento del cultivo de soja.
- El cultivo presentó mayor agresividad cuando se sembró a mayor densidad.
- No hubo diferencias en la competitividad del cultivo entre la siembra en grilla y en surcos.
- La practica de sembrar en grilla implica un gasto extra de combustible y tiempo que no se justifica ya que no aporta ninguna ventaja competitiva al cultivo.
Bibliografía
Aarssen L.W., Keogh T. 2002. Conundrums of competitive ability: what to measure? Oikos. 96: 531-542.
Brainard D. C., Bellinder R. R., DiTommaso A. 2005. Effects of canopy shade on the morphology, phenology, and seed characteristics of Powell amaranth (Amaranthus powellii).Weed Science. 53: 175-186.
Di Rienzo, J.A., Casanoves F., Balzarini M.G.,González L., Tablada M., Robledo C.W. 2013. InfoStat versión 2013. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina.
Ehleringer J. 1983. Ecophysiology of Amaranthus palmeri, a Sonoran desert summer annual. Oecologia.57:107–112.
Ehleringer, J. 1985. Annuals and perennials of warm deserts. Pages 162–180 in B.F. Chabot and H. A. Mooney, eds. Physiological Ecology of North American Plant Communities. New York: Chapman and Hall.
Ehleringer J., Forseth I. 1980. Solar tracking by plants. Science. 210 :1094–1098.
Goldberg D.E., Landa K. 1991. Competitive effect and response: hierarchies and correlated traits in the early stages of competition. The Journal of Ecology. 79: 1013-1030.
Grime J. 1979. Plant Strategies and vegetation process. John Wiley & Sons, New York. USA.
Heap, L. 2022.The international Survey of Herbicide Resistant seeds. Online. Internet. Disponible en : www.weedscience.org.
Keeley P.E., Carter C.H., Thullen R.J.1987. Influence of planting date on growth of Palmer amaranth (Amaranthus palmeri). Weed Science. 35:199–204.
Klingaman T. E., Oliver L. R. 1994. Palmer Amaranth (Amaranthus palmeri) Interference in Soybeans (Glycine max).Weed Science. 42: 523-527.
Massinga R. A., Currie R. S., Horak M. J., Boyer J. 2001. Interference of Palmer amaranth in corn. Weed Sci. 49:202–208.
Massinga, R. A., & Currie, R. S. (2002). Impact of Palmer amaranth (Amaranthus palmeri) on corn (Zea mays) grain yield and yield and quality of forage. Weed Technology, 16(3), 532-536.
McGilchrist C.A., Trenbath B.R.1971. A revised analysis of plant competition experiments. Biometrics. 27: 659-671.
Morichetti, S., Cantero, J. J., Núñez, C., Barboza, G. E., Ariza Espinar, L., Amuchastegui, A., & Ferrell, J. (2013). Sobre la presencia de Amaranthus palmeri (Amaranthaceae) en Argentina. Boletín de la Sociedad Argentina de Botánica, 48(2), 0-0.
Nurse R.E., DiTommaso A. 2005. Corn competition alters the germinability of velvetleaf (Abutilon theophrasti) seeds. Weed science. 53: 479-488.
REM. 2022. Red de conocimiento de malezas resistentes. Online. Internet. Disponible en: http://www.aapresid.org.ar/rem/mapas-rem/
Satorre E. H., Kruk B.C. 2016. Interacciones cultivo-maleza: competencia. Satorre E. H., Kruk B. C, de la Fuente E. B. Bases y herramientas para el manejo de malezas. Facultad de Agronomía. Universidad de Buenos Aires.140-178.
Satorre E.H., Snaydon R.W. 1992. A comparison of root and shoot competition between spring cereals and Avena fatua L. Weed Research 32: 45- 55.
Sauer, J. D. 1957. Recent migration and evolution of the dioecious amaranths. Evolution 11:11–31.
Snaydon R. W, y Satorre E.H. 1989. Modifications and interpretations of bivariate diagrams for plant competition data. Journal of applied Ecology. 26: 1043-1057.
Spitters C.J.T. 1983. An alternative approach to analysis of mixed cropping experiments. I. Estimation of competitive effects. Netherlands Journal of Agricultural Science 31: 1–11.
Tilman D. 1988. Mechanisms of competition for nutrients and light. En: Plant strategies and the dynamics and structure of plant communities. Princeton University Press, Princeton, New Jersey, USA.
Wang J. L., Klessig D. F., Berry J. O. 1992. Regulation of C4 gene expression in developing amaranth leaves. Plant Cell. 4:173–184.
Ward S. M., Webster T. M., Steckel. L.E.2013. Palmer Amaranth (Amaranthus palmeri): a review. Weed Technology.27 (1): 12-27.
Weiner J.1988. The influence of competition on plant reproduction. Doust J. L., Doust L. L. Ecology: Patterns and Strategies. New York: Oxford University Press. Eds. Plant Reproductive.228–245.
Weiner J., 2004. Allocation, plasticity and allometry in plants. Perspectives in Plant Ecology. Evolution and Systematics. 6, 207-215.
Wilson B. J.1988. Shoot competition and root competition. Journal of Applied Ecology. 25: 279-296.
Wilson S.D., Tilman D. 1995. Competitive responses of eight old field plant species in four environment. Ecology. 76: 1169-1180.

















