









Por: Fernando H. Oreja, Diego N. Donamaria, Elba B. de la Fuente – Catedra de Cultivos Industriales de la Facultad de Agronomía de la Universidad de Buenos Aires.
Introducción
Las malezas, junto con las plagas y las enfermedades, son algunas de las mayores preocupaciones para la producción agrícola en la Argentina y el mundo. Entendiendo por maleza a todo genotipo, población o especie vegetal que, por su abundancia y distribución interfiere en el tiempo y el espacio con las actividades humanas, generando algún efecto negativo de forma directa o indirecta. Dentro de los efectos negativos directos, el más importante es la caída del rendimiento de los cultivos debido a la competencia por recursos a lo largo del ciclo ontogénico. Por lo tanto, la magnitud de la merma en el rendimiento será producto del momento y grado de la interacción que se produzca.
En la Argentina, Digitaria sanguinalis más conocida como “pasto cuaresma”, “pata de gallina” o “pata de gallo”, tiene una ventana de emergencia muy amplia que puede ir desde inicios de la primavera hasta principios del verano. Esto genera la aparición de numerosas cohortes que le permiten escapar a los controles previos y posteriores a la siembra de los cultivos de verano. Esta característica, sumada a la gran producción de semillas, explican en parte su alta constancia en los cultivos.
Por otra parte, es una buena competidora por recursos debido al rápido crecimiento radical y aéreo (especie C4). Genera un tapiz herbáceo por medio de tallos rastreros o estolones que, sumado a la producción de sustancias alelopáticas, hacen de esta especie un serio problema en los cultivos estivales.
Para que las semillas del banco germinen, debe disminuir su nivel de dormición, para lo cual se requieren principalmente temperaturas frescas como las exploradas durante la estación invernal y humedad. Una vez que la dormición disminuye, la alternancia de temperaturas de primavera (entre 20°C y 30°C preferentemente) provoca la terminación de la dormición y favorece la germinación. Posteriormente, se produce la emergencia de las plántulas, regulada por la temperatura, la disponibilidad de agua y la profundidad de las semillas en el suelo. El estado de plántula es el momento crítico para lograr un control exitoso, evitando el crecimiento y la supervivencia de los individuos y la posterior producción de semillas que se suman al banco del suelo logrando perpetuar la especie a través del tiempo y el espacio.

Sembrar cultivos invernales, como trigo, avena o centeno, es una exitosa herramienta de manejo y control de malezas primavero – estivales, ya que reduce la abundancia de las mismas, suprimiendo o retardando su emergencia y reduciendo su incidencia en los cultivos de verano siguientes. Estos efectos de supresión se deben a que la presencia de un cultivo vivo genera una modificación en el ambiente que rodea a las semillas en el banco interfiriendo con el proceso germinativo. Un canopeo denso reduce la radiación fotosintéticamente activa (RFA) que llega al suelo, las temperaturas máximas y, por lo tanto, la amplitud térmica del suelo, limitando la terminación de la dormición de las semillas y la germinación. Por otra parte, se genera un cambio en la calidad de luz que llega al suelo, a través de la absorción de la luz roja por parte de las hojas, reduciendo la relación rojo/rojo lejano (R/RL) y condicionando la germinación de algunas especies.
Una vez que las semillas germinan y emergen las plántulas, los individuos dejan de crecer a expensas de las reservas provenientes de la planta madre y comienzan a requerir de recursos externos, compitiendo con sus vecinos por los recursos (luz, agua y nutrientes). Por ello, aumentos de la densidad del cultivo le otorgan una ventaja competitiva en la captación de los recursos esenciales, reduciendo los recursos disponibles para la maleza, afectando su crecimiento y fecundidad y reduciendo la cantidad de semillas que ingresan en el banco del suelo.
La emergencia anticipada del cultivo invernal con respecto a la de D. sanguinalis, permite llegar a la primavera con un tamaño de las plantas y volumen de biomasa aérea del cultivo lo suficientemente desarrollados como para generarle a la maleza un ambiente adverso para emerger y crecer satisfactoriamente.
“La introducción en la rotación de un cultivo invernal como el trigo es una opción exitosa para reducir la presencia de D. sanguinalis en el cultivo estival siguiente, no sólo en el corto plazo sino también a largo plazo”
Ya que el cultivo afectaría tanto la abundancia de las malezas en el cultivo siguiente (corto plazo) como su habilidad competitiva, su fecundidad y, por ello, la cantidad de semillas que ingresan al banco. Tanto los efectos sobre la germinación como sobre la competencia se verían más favorecida por cultivos de trigo sembrados a mayor densidad ya que el efecto sobre el ambiente debajo del canopeo y la competencia por recursos se intensifica más tempranamente en cultivos más densos.

El objetivo general del trabajo fue determinar los efectos de un cultivo de trigo sobre la dinámica poblacional de D. sanguinalis. Especialmente sobre los procesos de establecimiento, crecimiento, supervivencia y fecundidad de esta maleza.
Metodología
Se realizó un experimento en el campo experimental de la Cátedra de Cultivos Industriales de la Facultad de Agronomía de la Universidad de Buenos Aires (FAUBA), con un arreglo completamente aleatorizado. Los factores evaluados fueron la presencia del trigo con tres niveles de densidad de plantas, densidad normal (300 pl m-2), baja densidad (200 pl m-2) y sin cultivo (0 pl m-2) con cinco repeticiones.
La siembra del cultivo de trigo (variedad Don Mario ® Ceibo de ciclo corto) se llevó a cabo el día 13/07 con una sembradora experimental de siete líneas separadas a 17,5 cm. Dentro de cada parcela donde se registró presencia de D. sanguinalis en ciclos anteriores (evitando los surcos y entresurcos exteriores) se colocaron al azar marcos de alambre de 25cm x 25cm (625cm2 de superficie). Periódicamente se registró el número de plántulas emergidas de D. sanguinalis, marcando cada cohorte nueva (grupo de individuos que nacen al mismo tiempo) con alambres de diferentes colores en la base de cada planta y se eliminaban las plantas que no eran de la especie.

Al final del ciclo del cultivo se cosecharon las plantas identificando el estado fenológico alcanzado (plántula, adulto y/o adulto reproductivo). Con la cantidad de individuos que componían cada estado demográfico se determinaron las tasas demográficas de supervivencia y fecundidad para cada tratamiento. La tasa de supervivencia se obtuvo sumando el número de plantas adultas dividida la cantidad de plántulas emergidas para cada cohorte:
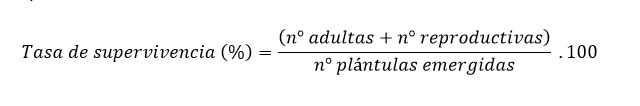
La tasa de fecundidad se calculó a partir del número de plantas que llegaban al estado reproductivo con inflorescencias emergidas fuera de la vaina, en relación a las plantas en estado adulto para cada cohorte:

Luego, se colocaron las plantas en una estufa a 60°C durante un período de 10 días (hasta peso constante), y posteriormente se pesaron para obtener así la biomasa aérea total para cada unidad experimental.
En intervalos de 15 a 20 días, durante días diáfanos y al mediodía, se registraron los datos ambientales de radiación incidente mediante un radiómetro (Cavadevices) y calidad de luz mediante un sensor rojo-rojo lejano (R-RL) (Skye). Para medir la radiación incidente se colocó la barra sobre la superficie del suelo de manera transversal a los surcos del cultivo (45º) y la calidad de luz se midió colocando el sensor en el entresurco sobre la superficie del suelo tomando los valores de tres orientaciones (arriba, izquierda y derecha) siempre mirando hacia el norte. A su vez, se midió la temperatura y humedad relativa del aire con “data loggers” colocados sobre sobre la superficie del suelo en los entresurcos.
Los efectos principales y sus interacciones se analizaron a través de un análisis de varianza (ANOVA) usando el programa estadístico InfoStat (versión 2020). Cuando el ANOVA fue significativo se utilizó el test de Tukey, con un nivel de significancia de p<0,05.
Resultados
A los 45 días desde la siembra (DDS), cuando el trigo se encontraba en estado Z2.2, los porcentajes de radiación incidente entre los tratamientos densidad normal y baja densidad no difirieron. A los 53 días, se observó una radiación incidente levemente mayor a baja densidad que a densidad normal. A inicios del mes de octubre, 81 días después de la siembra y con el trigo en estado de vaina engrosada (Z4), la radiación incidente se mantuvo en valores y diferencias similares entre ambos tratamientos siendo del 20,11% en densidad normal y 24,55% a baja densidad.
Luego de 95 DDS, con el trigo en estado de espigazón, (Z5) la radiación incidente aumentó levemente a 31,56% a densidad normal y 37.98% a baja densidad debido a la senescencia de las hojas basales del cultivo.
Finalmente, el 15 de noviembre (125 DDS) los porcentajes de radiación incidente fue similar entre ambos tratamientos. En las parcelas sin cultivo, la radiación incidente se mantuvo en valores de entre el 88% al 99% (Figura 1).

Figura 1: Evolución de la radiación incidente (%) durante el experimento, en cada uno de los tratamientos; sin cultivo (SC) (●), trigo baja densidad (TBD) (□), trigo alta densidad (TAD) (▲) en relación a los días después de la siembra (DDS).
En las parcelas sin trigo, las temperaturas promedio, tendieron a ser superiores a tratamientos con trigo a densidad normal y alta densidad entre los meses de septiembre y noviembre (50 y 100 días desde la emergencia, respectivamente).
A medida que el cultivo crecía y se desarrollaba e interceptaba radiación, la alternancia de temperaturas (diferencia entre las máximas y las mínimas) era mayor en las parcelas sin cultivo que en aquellas con cultivo, especialmente por mayores temperaturas máximas (Figura 2).

Figura 2. Temperatura diaria en las parcelas sin cultivo, con trigo a densidad normal y a baja densidad durante el experimento.
El canopeo del cultivo de trigo redujo la relación R/RL tanto en densidad normal como baja. Cuando el trigo presentaba dos macollos (Z2.2) a los 46 DDS, las parcelas sin cultivo mostraron valores de R/RL promedio de 0,88 contra 0,69 y 0,67 de parcelas de baja densidad y densidad normal, respectivamente. A los 83 DDS, en estado de vaina engrosada (Z4), la diferencia entre los tratamientos con y sin cultivo se fue ampliando, con valores de 0,80 para sin cultivo, 0,39 para la baja densidad y 0,35 a densidad normal.
Al final del experimento, a los 111 DDS, la relación R/RL disminuyó a 0,56, para el tratamiento sin cultivo. Tanto a baja densidad como a densidad normal de trigo no hubo diferencias en los valores de calidad de luz (Figura 3).

Figura 3: Evolución de la relación rojo: rojo lejano (R/RL) con sus desvíos, en función de los días desde la siembra del cultivo (DDS), en cada uno de los tratamientos; sin cultivo (SC) (●), trigo baja densidad (TBD) (□), trigo alta densidad (TAD) (▲). En el eje de abscisas se muestran los días después de la siembra (DDS) en que se registraron los datos (46, 53, 83 y 111 días).
Durante el experimento se registraron cuatro cohortes, la primera emergió el 4/9, la segunda el 19/9, la tercera el 16/10 y finalmente la cuarta el 1/11. La segunda cohorte fue la única donde en el tratamiento sin cultivo se observó una mayor cantidad de plántulas de la maleza, 707 m2 vs. 230 m2 y 300 m2 plántulas a densidad normal y baja respectivamente (un 38.5% y 32.9% menos de plántulas que sin cultivo). Entre los dos tratamientos con trigo no hubo diferencias significativas. No se observaron diferencias significativas (p<0,05) entre el total de plántulas por tratamiento, con medias de 1216 pl m-2 sin cultivo, 1238 pl m-2 a baja densidad y 860 pl m-2 a densidad normal.
En la segunda cohorte el número de plantas adultas fue mayor en el tratamiento sin cultivo (393 pl m-2) que en los tratamientos con densidad normal y baja densidad (96 y 60 pl m-2, respectivamente). En cambio, en la cuarta cohorte no hubo plantas adultas en el tratamiento sin cultivo y sí con cultivos (35 y 25 pl m-2) en alta y baja densidad, respectivamente). En las otras dos cohortes no hubo diferencias entre tratamientos (Figura 4).
Si bien las tasas de supervivencia no difirieron significativamente entre tratamientos por la alta dispersión de los datos, se observó que el tratamiento sin cultivo en las cohortes 1, 2 y 3, tendió a tener una mayor tasa de supervivencia que los tratamientos con cultivo, independientemente de la densidad del cultivo (Figura 4).
Sólo en las parcelas sin cultivo hubo plantas que alcanzaron el estado reproductivo, independientemente de las cohortes 1, 2 o 3. En la cohorte 4 no se observaron individuos en estadios reproductivos en ninguno de los tres tratamientos al momento en que el cultivo de trigo se encontraba en madurez de cosecha. Si bien no se observaron diferencias significativas, la cohorte 2 tendió a una mayor cantidad de adultos reproductivos (Figura 4).



Figura 4. Tabla de vida de Digitaria sanguinalis sin cultivo de trigo, creciendo con un cultivo de trigo a baja densidad (200 pl m-2) y con un cultivo de trigo a densidad normal (300 pl m-2). Los valores del número de plántulas, plantas adultas y reproductivas corresponden al valor promedio de cada marco (625cm2). Las tasas demográficas están expresadas en porcentaje. Letras distintas indican diferencias significativas entre variables de la misma cohorte para los diferentes tratamientos según Tukey (p<0,05).
Conclusiones
La presencia del cultivo de trigo:
- Modificó el ambiente en el que se encontraban las semillas de D. sanguinalis independientemente de la densidad (baja o normal). En particular, disminuyó la radiación incidente con lo cual se redujeron las temperaturas máximas y la alternancia de temperaturas y la fracción RL de la luz que llega al suelo, con lo cual se redujo la relación R/RL.
- Afectó la germinación-emergencia de las semillas de D. sanguinalis de la segunda cohorte, a través de una menor alternancia de temperaturas y una menor relación R/RL, ambos requerimientos para la terminación de la dormición de esta especie.
- Redujo la disponibilidad de la radiación incidente para D. sanguinalis, lo que redujo la supervivencia de las plántulas y la fecundidad de la segunda cohorte, principalmente.
- Generó un ambiente desfavorable para la maleza de manera que no hubo recursos suficientes para que las plantas alcanzaran el estado reproductivo al momento de la cosecha del cultivo.


















